:: Celulas
Vamos entrar agora no universo microscópico
das células. Para entendermos a sua natureza, vamos
estudá-las sob diversos aspectos, mas tendo sempre
em mente que o importante é a compreensão da
célula como um todo vivo e não uma máquina
dividida em setores. As células possuem uma grande
diversidade de origens, formas, tamanhos e funções.
Esses aspectos representam variações de um mesmo
tema, pois muitas estruturas e funções se repetem
em todos os tipos celulares. De início, precisaremos
conhecer aspectos, como tamanho, forma, volume, relação
entre volume e superfície e, também, a durabilidade
dos ciclos celulares. Existem dois modelos estruturais de
células: o procarionte e a eucarionte, sendo este último
do tipo animal e do tipo vegetal. Elas constituem-se em verdadeiras
unidades básicas, morfológicas e fisiológicas
dotadas de incrível dinâmica e onde a vida se
manifesta de forma independente e ativa. Portanto, veremos
que as células, como entidades vivas, são dotadas
de uma complexidade estrutural e funcional superior, permitindo-lhes
uma infinidade de capacidades e transformações
que são próprias da vida. Afinal, ainda não
se conhece vida sem células. As células são,
em sua grande maioria, menores do que a capacidade de resolução
do olho humano, portanto só podem ser observadas com
o uso de microscópios. A unidade de medida mais comumente
empregada para mensurar as células é o mícron,
que corresponde à milésima parte do milímetro
(1mm : 1000). A maior parte das células animais e vegetais
oscila entre 10 e 50 mícrons. Entretanto, podemos encontrar
células que fogem desse padrão mediano de tamanho.
Uma bactéria esférica, o micoplasma ou PPLO,
causadora de uma infecção respiratória
em bovinos, é considerada a menor célula conhecida,
com apenas 0,1 mícron de diâmetro. Devido a essa
dimensão, só é visível ao microscópio
eletrônico. Em oposição aos micoplasmas,
podemos citar a alga verde Acetabularia que possui forma de
um pequeno guarda-chuva, sendo 10. 000 vezes maior que os
micoplasmas, portanto visível a olho nu. No homem,
os prolongamentos das células nervosas que saem da
coluna vertebral e vão até os pés podem
atingir até 1 metro de comprimento. Nos vegetais, algumas
fibras dos tecidos de sustentação podem atingir
alguns centímetros. As células são dotadas
de grande dinamismo e apresentam formas extremamente variáveis.
A grande maioria das células possui forma constante
(cúbica, esférica, prismática, estrelada,
ramificada, fusiforme e outras), porém algumas modificam
continuamente sua forma sendo denominadas polimorfas, como
as amebas, os leucócitos, macrófagos e outras.
Vários são os fatores que influenciam a determinação
da forma celular. Dentre eles destacamos a tensão superficial
da membrana plasmática, o nível de viscosidade
do citoplasma, o meio em que estão inseridas as células,
a pressão exercida por células vizinhas, as
informações genéticas das células,
entre outras. Normalmente, a forma das células dos
animais e vegetais é condicionada pela função
que desempenham no organismo. As células epiteliais
de revestimento, por exemplo, são achatadas e poliédricas
de modo que possam cobrir a maior superfície possível,
encaixando-se perfeitamente umas nas outras, não deixando
espaços e cumprindo bem as funções de
revestimento e proteção. As células musculares
são alongadas, pois são adaptadas à contração,
portanto não teria sentido se fossem cúbicas
ou esféricas para cumprir tal função.
Lei de Driesch
A Lei de Driesch ou Lei do Volume Celular
Constante diz que "o volume celular é constante
nas células de um mesmo tecido em indivíduos
de mesma espécie e em mesma fase de desenvolvimento,
não interessando o tamanho dos indivíduos".
Portanto, o que o diferencia dos seus colegas em tamanho é
o número de células que possuem e não
o volume que apresentam. Seus colegas maiores possuem maior
quantidade de células, e o contrário se aplica
para os colegas menores. As células constituintes do
tecido nervoso e do tecido muscular estriado não são
subordinadas a essa lei. As células nervosas podem
possuir prolongamentos que saem da cabeça e vão
até os pés. Então essas células
serão mais volumosas em um indivíduo com 1,80m
do que em um de 1,50m. As células musculares, quando
continuamente estimuladas, podem sofrer hipertrofia aumentando
o seu volume. Isso pode ser observado nos halterofilistas
que apresentam musculatura bastante desenvolvida em função
da continua atividade.
Lei de Spencer
Para uma melhor compreensão
dessa relação, vamos observar a Lei de Spencer,
segundo a qual "a superfície celular varia com
o quadrado da dimensão linear, e o volume com o cubo
da mesma dimensão". As células estão
continuamente trocando substâncias entre si e com o
meio em que se encontram. Os nutrientes de que uma célula
precisa, penetram no seu interior, passando pela membrana
(superfície), sendo então aproveitados no citoplasma
(volume). Isso faz com que a célula cresça,
aumentando o citoplasma e a membrana (volume e superfície).
Quando essa relação é rompida e o volume
citoplasmático passa a crescer mais que a superfície
da membrana, a célula buscará equilibrar-se
novamente, dividindo-se em duas novas células. Portanto,
o desequilíbrio entre o volume e a superfície
pode ser um fator indutor da divisão celular.
:: CICLO VITAL DAS CÉLULAS
A longevididade de uma célula
é muito variável conforme a espécie.
No organismo humano, há células que duram muitos
anos. Algumas têm a sua duração contada
em dias. Outras acompanham o indivíduo pot toda a vida,
do nascimento à morte. Sob esse ponto de vistas, as
células são classificadas em lábeis,
estáveis e permanentes.
• Células lábeis:
são células
de curta duração. De modo geral, não
se agrupam de forma fixa na organização dos
tescidos, não se reproduzem e resultam de diferenciação
rápida de células indiferenciadas de origem
embrionária. Como tal , se classificam os gametas (duram
dois ou três dias) e as hemácias ou glóbulos
vermelhos do sangue (no máximo 120 dias).
• Células estáveis:
constituem a grande maioria dentre as numerosas variedades
celulares do nosso organismo. São células que
se diferenciam durante o desenvolvimento embrionário
e depois mantêm um ritmo constante de multiplicação.
Assim ocorre com as fibras musculares lisas e os diversos
tipos de células epiteliais e conjuntivas. Podem duram
meses ou anos. As células dos vegetais também
se classificam nesse grupo.
• Células permanentes:
resultam de uma diferenciação celular muito
precoce no embrião. Duram toda a vida. Atingem alto
grau de especialização. Por isso, depois de
concluída a formação embrionária,
perdem a capacidade de reprodução. É
o que se verifica com as fibras musculares estriadas e com
os neurônios. Não há renovação
dessas células nos organismo depois do nascimento.
Por isso mesmo, são inviáveis os transplantes
de coração.
:: CÉLULAS PROCARIOTAS
Partindo-se do princípio estudado
sobre a origem da vida, de que o mais simples gerou o mais
complexo, os cientistas em geral acreditam que as células
procariotas tenham sido as primeiras a surgirem em nosso planeta.
As bactérias e as cianofíceas, ou algas azuis,
são seres unicelulares, isolados ou coloniais, que
ocupam os mais variados ambientes. As bactérias e as
cianofíceas também são os únicos
seres vivos que apresentam uma estrutura celular procarionte.
Por isso, estão classificados no Reino Monera que lhes
é exclusivo. Se analisarmos uma célula procariótica,
perceberemos a sua simplicidade estrutural, o material nuclear
não fica delimitado por uma membrana, constituindo
um núcleo indefinido. Nos seres procariontes, o material
nuclear fica espalhado ou disperso pelo citoplasma. Muitas
células procariotas possuem uma extraordinária
capacidade de adaptação às condições
ambientais desfavoráveis. O papel biológico
desses pequenos organismos é fundamental para a saúde
da biosfera, pois são colonizadores de regiões
abióticas, decompositores de matéria orgânica
e controlam populações de outros seres vivos.
As cianofíceas ou algas azuis realizam a fotossíntese,
por isso apresentam uma estrutura um pouco mais sofisticada
que a das bactérias; porém, assim como as bactérias,
não possuem um núcleo definido, sendo, portanto,
procariontes.
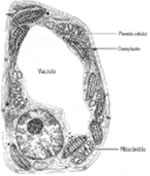
:: CÉLULA EUCARIOTAS
Os cientistas acreditam que
as células procariotas tenham, por especialização
e necessidade de sobrevivência, originado as células
eucariotas. Assim, as células eucariotas são
mais evoluídas e mais complexas estruturalmente. Isso
se comprova pela riqueza de membranas e compartimentos internos,
além de organelas e inclusões presentes no citoplasma.
Entre as várias membranas internas, podemos citar a
carioteca ou membrana nuclear, que abriga no seu interior
a maior parte do material genético da célula,
constituindo assim um núcleo definido. Os animais,
vegetais, fungos e protistas são constituídos
por células eucariotas. Porém, também
apresentam diferenças entre si. Essas diferenças
são especialmente evidentes entre células de
animais e vegetais. As células dos animais apresentam
o glicocálix sobre a face externa da membrana plasmática,
além de lisossomos. As células vegetais, por
realizarem a fotossíntese, necessitam de um equipamento
celular especial. Possuem um conjunto de plastídeos
específicos para a fotossíntese, além
de uma parede celular externa de natureza celulósica
e grandes vacúolos de suco celular.
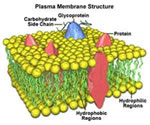
:: MEMBRANA PLASMÁTICA
Você conhece o fato de os países
possuírem um determinado território, o qual
é delimitado por uma fronteira. Desse modo, estabelece-se
um limite físico ou uma individualidade para esse território.
Teoricamente, através dessa fronteira, pode-se estabelecer
um controle do de entrada e saída de pessoas e produtos,
pois as nações, assim como as células,
não podem existir como sistemas fechados. Apesar de
as fronteiras entre países serem imaginárias,
podemos usar esta comparação para entendermos
um pouco sobre a membrana plasmática, já que
a mesma membrana representa um limite físico da célula.
Esta membrana mantém o conteúdo interno ou citoplasmático
separado, porém não isolado do meio externo,
visto que ela possui uma capacidade seletiva quanto ao que
entra e sai das células. As membranas celulares são
muito delgadas, por exemplo: a membrana plasmática,
que mede cerca de 75 Å de espessura, devido a essa dimensão,
não são visíveis ao microscópio
óptico comum. Os biologistas suspeitavam da existência
de uma membrana celular,pois as células quando rompidas
modificavam a composição química do meio
em que se encontravam. Portanto, deveria existir algo que
mantivesse sua individualidade. Apenas neste século,
com a utilização de técnicas de microscopia
eletrônica, é que foi possível estudá-la
adequadamente
A Estrutura da Membrana Plasmática
Para começarmos a estudar a
estrutura da membrana plasmática, é necessário
conhecermos um pouco de sua composição química.
Os lipídios e as proteínas são exemplos
de compostos que participam diretamente na formação
de membranas celulares. Na verdade, são os principais
componentes estruturais de membranas celulares com os lipídios,
principalmente os fosfolipídios, totalizando 25% a
40% do total e as proteínas 60% a 75%. Em menor quantidade,
podem-se encontrar antígenos, enzimas, glicídios
e outras moléculas permanentes ou transitórias.
Por isso, as membranas celulares são denominadas lipoprotéicas,
pois representam uma associação entre lipídios
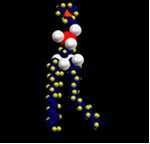
e proteínas. Em 1954, sabendo-se que uma molécula
fosfolipídica da membrana apresenta um pólo
de afinidade com a água (hidrófilo) e outro
pólo de aversão à água (hidrófobo),
os cientistas Dawson e Danielli propuseram um modelo segundo
o qual a membrana plasmática seria formada por duas
camadas de lipídios revestidas por proteínas.
| |
|
 |
As moléculas de lipídios
se arranjariam, de modo que os pólos hidrófobos
ficariam no interior da membrana confrontando-se, enquanto
os pólos hidrófilos ficariam na parte externa
com uma face em contato com o meio intracelular e outra com
o meio extracelular. Os pólos hidrófilos estariam
recobertos por uma camada fixa de proteínas que se
encaixaria nas camadas lipídicas. O modelo admitia
a presença de poros para justificar a comunicação
da célula com o meio externo. No entanto, o modelo
não era satisfatório para explicar todos os
aspectos relacionados às necessidades das células
e às capacidades da membrana, em especial, a de selecionar
o trânsito de substâncias entre a célula
e o meio em que ela se encontra. Tal capacidade é denominada
de permeabilidade seletiva e é característica
de todas as membranas celulares. O modelo estrutural mais
aceito atualmente é o proposto pelos cientistas Singer
e Nicholson em 1972, e corresponde a um aprimoramento do modelo
anterior de Dawson e Danielli. Segundo o modelo de Singer
e Nicholson, a bicamada lipídica está presente,
como no modelo proposto por Dawson e Danielli, porém
as proteínas não estão imersas na bicamada
revestindo-a interna e externamente. Na verdade, as proteínas
apresentam uma mobilidade especial, podendo se deslocar lateralmente
ou atravessar a bicamada lipídica, projetando-se nas
superfícies interna ou externa da membrana plasmática.
Conclui-se, portanto, que a membrana é relativamente
fluida, pois as moléculas de proteínas apresentam
certa liberdade de movimentação. Por isso, o
modelo de Singer e Nicholson é denominado mosaico fluido.
Esse modelo é satisfatório para que as membranas
celulares possam exercer todas as suas capacidades e funções,
especialmente a sua permeabilidade seletiva. Ao microscópio
eletrônico, a membrana plasmática apresenta um
aspecto trilaminar característico. São duas
lâminas laterais mais densas, correspondendo aos pólos
hidrófilos dos lipídios mais as proteínas,
e uma lâmina central mais clara, que corresponde aos
pólos hidrofóbicos da bicamada lipídica.
Este aspecto se repete em todas as membranas da célula,
tanto na membrana plasmática quanto na de organelas,
como mitocôndrias, cloroplastos, retículo endoplasmático
e outros. Diante desse fato, o cientista Robertson criou,
no final da década de 50, o termo unidade de membrana,
referindo-se ao fato de todas as membranas celulares apresentarem
uma estrutura similar.
:: REFORÇOS DA MEMBRANA
Em células de vegetais, células
animais, bactérias e de outros seres, pode-se encontrar
a superfície externa da membrana com uma série
de envoltórios com composição e funções
variadas. A parede bacteriana é um envoltório
externo de muitas bactérias e tem como principal função
a manutenção da forma específica de cada
bactéria, protegendo-as das variações
osmóticas do meio. A parede celular é o envoltório
característico das células vegetais. Ela possui
uma composição química complexa e atua
principalmente como estrutura esquelética.
O glicocálix é um envoltório
externo de células animais formado por glicolipídios
e glicoproteínas. Além da função
de adesão, o glicocálix tem uma importante função
de reconhecimento do meio. Cada célula tem o seu glicocálix
característico como uma espécie de "impressão
digital da célula". Esta característica
permite que as células de um determinado tecido reconheçam-se
entre si, limitando o seu crescimento por inibição
de contato. As células cancerosas apresentam o seu
glicocálix alterado, por isso não param o seu
crescimento pelo contato com as células vizinhas, formando
então massas celulares que se constituem nos tumores.
:: TRANSPORTE DE MEMBRANA
Transporte através da membrana:
Os componentes hidrofóbicos, solúveis nos lipídios,
atravessam facilmente a membrana, por ser esta constituída
de uma bicamada lipídica , como é o caso dos
ácidos graxos, hormônios esteróides e
anestésicos. As substâncias hidrófilas,
insolúveis nos lipídios, penetram nas células
com mais dificuldade, dependendo do tamanho da molécula
e também de suas características químicas.
A configuração molecular poderá permitir
que a substância seja transportada por intermédio
de um dos mecanismos especiais desenvolvidos durante a evolução,
como o transporte ativo e a difusão facilitada.
Difusão Passiva
Muitas substâncias penetram nas
células ou delas saem por difusão passiva, isto
é, como a distribuição do soluto tende
a ser uniforme em todos os pontos do solvente, o soluto penetra
na célula quando sua concentração é
menor no interior celular do que no meio externo, e sai da
célula no caso contrário. Neste processo não
há consumo de energia.
Difusão Facilitada
Algumas substâncias, como a glicose,
galactose e alguns aminoácidos têm tamanho superior
a 8 Angstrons, o que impede a sua passagem através
dos poros. São, ainda, substâncias não
solúveis em lipídios, o que também impede
a sua difusão pela matriz lipídica da membrana.
No entanto, estas substâncias passam através
da matriz, por transporte passivo, contando, para isto, com
o trabalho de proteínas carreadoras (proteínas
transportadoras). A combinação entre a glicose,
por exemplo, e a proteína carreadora forma uma combinação
lipossolúvel que passa, então, a difundir-se
de um lado para outro da membrana. Do outro lado da membrana,
a glicose separa-se do carreador, passa para o interior da
célula, enquanto o carreador retorna ao meio externo
para buscar mais moléculas de glicose. Este processo
está esquematizado abaixo:
A velocidade com que a difusão
facilitada acontece depende da diferença de concentração
de substâncias nos dois lados da membrana, da quantidade
de carreadores disponíveis e da velocidade com que
as reações se processam. No caso da glicose,
a velocidade de sua difusão é grandemente aumentada
com a presença de maior quantidade de insulina, hormônio
secretado pelo pâncreas. Não se sabe, ainda,
se o efeito da insulina está no aumento dos carreadores
ou no aumento da velocidade de processamento das reações
químicas entre a glicose e o carreador.
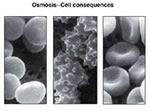
Osmose
É um fenômeno de difusão
em presença de uma membrana semipermeável. Nele,
duas soluções de concentrações
diferentes estão separadas por uma membrana que é
permeável ao solvente e praticamente insolúvel
ao soluto. Há, então, passagem do solvente de
onde está em maior quantidade (solução
hipotônica) para onde está em menor quantidade
(solução hipertônica).
Com esta passagem, verifica-se um
aumento da quantidade de água na solução
hipertônica, fazendo com que haja maior diluição
da solução e, consequentemente, diminuição
da sua concentração. Podemos dizer que é
a osmose que possibilita isotonia entre uma substância
hipertônica e uma hipotônica, com passagem de
solvente através de uma membrana semipermeável.
Essa pode, inclusive,ser fatal para a célula, como
no caso da hemácias que, em presença de soluções
pouco concentradas, sofrem hemólise. A presença
da parede celular nas células vegetais torna peculiar
este fenômeno, onde a célula vegetal, mesmo em
meios muito pouco concentrados em relação aos
seus vacúolos, não explode ( deplasmólise).
Em soluções hipertônicas esses dois tipos
de células apresentam, respectivamente, crenação
e plasmólise.
Transporte Ativo
Neste caso há consumo de energia
e a substância pode ser transportada de um local de
baixa concentração para um outro de alta concentração.
O soluto na difusão ativa pode ser transportado contra
um gradiente. O transporte ativo é bloqueado pelos
inibidores da respiração como o dinitrofenol,
cianetos, azida, e iodoacetato, inibidores da síntese
de ATP.
Transporte em Massa
O transporte em quantidade para dentro
da célula, também chamado endocitose, é
feito por dois processos denominados fagocitose e pinocitose.
Quando a transferência de macromoléculas tem
lugar em sentido inverso, isto é, do citoplasma para
o meio extracelular, o processo recebe o nome genérico
de exocitose.
- Fagocitose:
É o nome dado ao processo pelo
qual a célula, graças à formação
de pseudópodos, engloba, no seu citoplasma, partículas
sólidas. A fagocitose é um processo seletivo,
conforme pode ser observado no exemplo da fagocitose de paramécios
pelas amebas. Nos mamíferos, a fagocitose é
feita por células especializadas na defesa do organismo,
como os macrófagos.
- Pinocitose:
É o nome dado ao processo pelo
qual a célula, graças à delgadas expansões
do citoplasma, engloba gotículas de líquido.
Formam-se assim vacúolos contendo líquido, que
se aprofundam no citoplasma tornando-se cada vez menores,
o que sugere uma transferência de líquido para
o hialoplasma. Muitas células exibem esse fenômeno,
como os macrófagos e as células endoteliais
dos capilares sangüíneos. No processo da pinocitose
formam-se longas projeções laminares da superfície
celular, visíveis ao microscópio óptico,
que dão origem a vesículas também grandes
no processo chamado de macropinocitose. A micropinocitose
é de ocorrência mais geral e dá origem
a vesículas menores, visíveis somente no microscópio
eletrônico.
:: ESPECIALIZAÇÕES DA
MEMBRANA PLASMÁTICA
- Interdigitações:
São saliências
e reentrâncias da membrana celular que se encaixam em
estruturas complementares das células vizinhas.
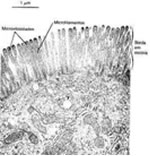
- Microvilosidades:
São especializações
apicais da membrana. Elas estão presentes na superfície
livre das células do intestino delgado, responsáveis
pela absorção de nutrientes. Cada célula
intestinal deste tipo possui em média três mil
microvilosidades. Em 1 mm2 de superfície intestinal,
existem cerca de 200 000 dessas especializações.
Elas são evaginações permanentes da membrana
com o aspecto digitiforme, que ampliam consideravelmente a
superfície de contato da célula com os nutrientes
vindos da digestão, para melhorar assim a função
de absorção intestinal.
- Desmosomos:
Cada desmosomo tem a forma de uma placa
arredondada e é constituído pelas membranas
de duas células vizinhas. Devido à função
de adesão e à sua distribuição
descontínua, o desmosomo é também chamado
de macula adherens.
- Zônula de Adesão:
É uma formação
encontrada em certos epitélios de revestimento, circundando
a parte apical das células. Sua estrutura é
semelhante à dos desmosomas, porém a zônula
de adesão é um cinto contínuo em volta
da célula. As suas funções são
promover a adesão entre as células e oferecer
local de apoio para os filamentos que penetram nos microvilos
das células epiteliais com orla em escova.
- Zônula Oclusiva:
É uma faixa contínua
em torno da zona apical de certas células epiteliais
que veda completamente o trânsito de material por entre
as células. Outra função da zônula
oclusiva, também chamada junção oclusiva,
é permitir a existência de potenciais elétricos
diferentes, conseqüência de diferenças na
concentração iônica entre as duas faces
da lâmina epitelial.
- Complexo Juncional:
Está presente em vários
epitélios próximo à extremidade celular
livre, sendo constituído dos seguintes elementos: zônula
oclusiva, zônula de adesão e uma fileira de desmosomas.
O complexo juncional é uma estrutura de adesão
e vedação.
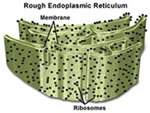
Retículo Endoplasmático
É uma rede de estruturas tubulares
e vesiculares achatadas, sendo que os túbulos e as
vesículas são interconectados uns aos outros.
Por outro lado, suas paredes são formadas por membranas
de bicamadas lipídicas, contendo grandes quantidades
de proteínas, de forma semelhante à membrana
celular. Podemos distinguir dois tipos de retículo
endoplasmático: o retículo endoplasmático
rugoso ou granular (RER) e o retículo endoplasmáticos
liso ou agranular (REL). O RER também é chamado
de ergastoplasma e é formado por sacos achatados, cujas
membranas têm aspecto rugoso devido à presença
de grânulos (ribossomos) aderidos à sua superfície
externa, voltada para o citosol. O REL é formado por
estruturas membranosas tubulares, sem ribossomos aderidos
e, portanto, de superfície lisa.
Funções do Retículo
Endoplasmático:
- Produção de
Lipídios: A
lecitina e o colesterol são exemplos de componentes
lipídicos que existem em todas as membranas celulares
e são produzidas no REL. Outros tipos de lipídios
produzidos são os hormônios esteróides,
dentre os quais estão a testosterona e o estrógeno
(hormônios sexuais produzidos nas células das
gônadas de animais vertebrados);
- Desintoxicação:
O REL participa dos
processos de desintoxicação do organismo, sendo
que nas células do fígado as substâncias
tóxicas são absorvidas e posteriormente são
modificadas ou destruídas, de modo a não causarem
danos ao organismo. A atuação do retículo
das células hepáticas permitem eliminar parte
do álcool, medicamentos e outras substâncias
potencialmente nocivas que ingerimos.
- Armazenamento de Substâncias:
Os vacúolos das células vegetais são
exemplos de bolsas membranosas derivadas do REL que crescem
pelo acúmulo de soluções aquosas ali
armazenadas.
- Produção de
Proteínas: As
proteínas fabricadas no RER (devido à presença
dos ribossomos) penetram nas bolsas e se deslocam em direção
ao aparelho de Golgi, passando pelos estreitos e tortuosos
canais do REL.
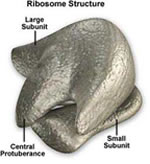
Ribossomos
Ribossomos são os locais de
síntese de proteína. Eles não são
limitados por membranas e portanto ocorrem tanto em procariontes
quanto em eucariontes. Os Ribossomos de Eucariontes são
ligeiramente maiores que os de procariontes. Estruturalmente,
o ribossomo consiste em uma sub-unidade pequena e outra maior.
Bioquimicamente o ribossomo consiste em RNA ribossômico
(RNAr) e umas 50 proteínas estruturais. Freqüentemente
os ribossomos crescem em cachos no retículo de endoplasmático,
eles se assemelham a uma série de fábricas que
juntam formando algo parecido com uma via férrea.
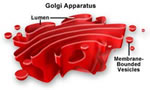
Complexo de Golgi
O complexo de Golgi está presente
em praticamente todas as células eucariontes e em geral
é formado por quatro ou mais camadas empilhadas de
delgadas vesículas achatadas chamadas de dictiossomo,
que situam-se próximas ao núcleo.
Funções do Complexo
de Golgi:
- Secreção
de Enzimas Digestivas: As
enzimas digestivas do pâncreas são exemplo de enzimas
produzidas no RER e levadas até as bolsas do complexo
de Golgi, onde são empacotadas em pequenas bolsas, que
se desprendem dos dictiossomos e se acumulam em um dos pólos
da célula pancreática. A produção
de enzimas digestivas pelo pâncreas é apenas um
entre muitos exemplos do papel do complexo de Golgi nos processos
de secreção celular.
-
Formação do Acrossomo do Espermatozóide:
O acrossomo é uma bolsa de enzimas
digestivas do espermatozóide maduro, que irão
perfurar as membranas do óvulo e permitir a fecundação.
- Formação
da Lamela Média em Células Vegetais:
A lamela média é a primeira
membrana que separa duas células recém - originadas
da divisão celular. Os dictiossomos acumulam o polissacarídio
pectina, que é eliminado entre as células –
irmãs recém – formadas, constituindo a primeira
separação entre elas e, mais tarde, a lâmina
que as mantém unidas.
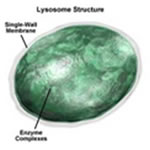
Lisossomos
Os lisossomos são bolsas circundadas
por típica membrana de bicamada lipídica e cheias
com grande número de pequenos grânulos, que são
agregados protéicos de enzimas hidrolíticas (digestivas)
capazes de digerir diversas substâncias orgânicas.
São originados no complexo de Golgi e estão presentes
em praticamente todas as células eucariontes.
Tipos de Lisossomos:
- Lisossomo Primário:
É o lisossomo propriamente dito, ou seja, a vesícula
possuindo no seu interior as enzimas digestivas.
- Lisossomo Secundário:
Denomina-se também de Vacúolo Digestivo e resulta
da fusão do lisossomo primário com a partícula
englobada.
- Corpúsculo Residual:
É a vesícula lisossômica que por exocitose
elimina na periferia celular o material não assimilado.
- Vacúolo Autofágico:
É quando a vesícula lisossômica digere uma
partícula pertencente à própria célula.
A autofagia é uma atividade indispensável à
sobrevivência da célula.
Funções dos
Lisossomos:
-
Digestão Intracelular:
A digestão ocorrerá no interior dos vacúolos
digestivos, que são bolsas originadas pela fusão
do lisossomo com o fagossomo ou pinossomo e contêm partículas
capturadas do meio externo.A digestão intracelular pode
ser classificada em: Autofagia – quando os lisossomos
digerem uma partícula pertencente à própria
célula e Heterofagia – quando a partícula
digerida pelos lisossomos é proveniente do meio extracelular.
- Desaparecimento da Cauda do Girino: A regressão da
cauda dos girinos se dá pela autodestruição
de células pelas enzimas lisossômicas. O material
resultante da autodigestão da cauda entra na circulação
sangüínea e é reutilizado.
- Autofagia Pós –
Morte
Vacúolos
Qualquer pedaço no citoplasma delimitado por um pedaço
de membrana lipoprotéica. As variedades mais comuns são:
" Vacúolos relacionados com a digestão intracelular
" vacúolos contráteis (ou pulsáteis)
" vacúolos vegetais As inclusões São
formações não vivas existentes no citoplasma,
como grãos de amido gotas de óleo. O conjunto
de inclusões denomina-se paraplasma. A seqüência
das estruturas formadas durante a digestão intracelular
é: Vacúolo alimentar, Vacúolo digestivo
e Vacúolo residual.
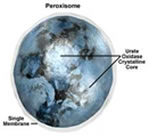
Peroxissomos
Os peroxissomos são, em termos físicos, semelhantes
aos lisossomos, mas diferem em dois aspectos importantes: Primeiro
acredita-se que sejam formados por auto – replicação
(ou talvez por brotamento do REL) e não pelo complexo
de Golgi; Segundo que eles contêm oxidases e não
hidrolases. Além de conterem enzimas que degradam gorduras
e aminoácidos, têm também grandes quantidades
da enzima catalase, que converte o peróxido de hidrogênio
( água oxigenada ) em água e gás oxigênio.
Os peroxissomos estão presentes
em grandes quantidades nas células de defesa como os
macrófagos e também existem nas células
vegetais, onde participam do processo da fotorespiração.
A função dos peroxissomos no metabolismo celular
ainda é pouco conhecida, mas acredita-se que participem
dos processos de desintoxicação da célula.
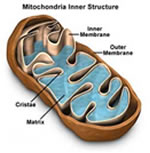
Mitocôndrias (Condrioma)
As mitocôndrias
são formadas principalmente por duas bicamadas lipídicas:
uma membrana externa e outra membrana interna. Enquanto a membrana
externa é lisa, a membrana interna possui inúmeras
pregas chamadas cristas mitocondriais, nas quais se fixam enzimas
oxidativas. A cavidade interna das mitocôndrias é
preenchida por um fluido denominado matriz mitocondrial contendo
grande quantidade de enzimas dissolvidas, necessárias
para a extração de energia dos nutrientes. As
mitocôndrias são verdadeiras “casas de força”
das células, pois produzem energia para todas as atividades
celulares. Sua composição química é
riquíssima, notando-se principalmente a presença
de DNA, RNA, proteínas, carboidratos, enzimas, ATP (adenosina
– trifosfato), ADP (adenosina – difosfato), etc.
São encontradas nas células eucariontes, sendo
substituídas pelos mesossomos nas bactérias. No
interior das mitocôndrias ocorre a respiração
celular, que é o processo em que moléculas orgânicas
de alimento reagem com gás oxigênio, transformando
– se em gás carbônico e água e liberando
energia. Toda mitocôndria surge da reprodução
de uma outra mitocôndria, sendo que a divisão da
mitocôndria denomina-se Condrocinese ou Condrogênese.
Funções
da Mitocôndria:
- Produção de Energia;
- Respiração Celular através do Ciclo
de Krebs e da Cadeia Respiratória.
Origem das Mitocôndrias:
Durante os anos oitenta, Lynn Margulis propôs a teoria
da endosimbiose para explicar a origem das mitocôndrias
e cloroplastos de procariontes. De acordo com esta idéia,
um procarionte maior engolfou ou cercou um procarionte menor
há uns 1.5 bilhão ou 700 milhões de anos
atrás.Em vez de digerir o organismo menor, o grande
e o pequeno entraram em um tipo de simbiose conhecido como
mutualismo, em que ambos os organismos se beneficiam e nenhum
é danificado. O organismo maior ganhou excesso de ATP
fornecido pela "protomitocôndria" e açúcar
em excesso fornecidos pelo " protocloroplasto ",
enquanto fornecia um ambiente estável e as matérias-primas
que o endosimbionte requeria. Esta relação é
tão forte que agora células de eucarionte não
podem sobreviver sem mitocôndria (igualmente eucariontes
fotossintéticos não podem sobreviver sem cloroplastos),
e os endosimbiontes não podem sobreviver fora dos anfitriões.
Quase todos eucariontes têm mitocôndria.
Plastos
Os plastos são orgânulos citoplasmáticos
encontrados nas células de plantas e de algas. São
classificados em:
- Cromoplastos:
São plastos coloridos que armazenam pigmentos.
Plastos |
Pigmentos |
Cor |
| Cloroplastos |
Clorofila |
Verde |
| Xantoplastos |
Xantofila |
Amarelo |
| Eritroplastos |
Eritrofila |
Vermelho |
| Cianoplastos |
Cianofila |
Azul |
| Feoplastos |
Feofila |
Parda |
- Leucoplastos
: São plastos incolores que armazenam
substâncias nutritivas como os Amiloplastos (amido),
os Oleoplastos (óleos) e os Proteoplastos (proteínas).
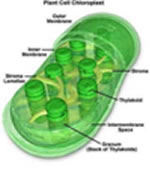
Os cloroplastos são orgânulos citoplasmáticos
discóides que apresentam duas membranas envolventes
e inúmeras membranas internas, que formam pequenas
bolsas discoidais e achatadas chamadas tilacóides.
Os tilacóides se organizam uns sobre os outros e formam
estruturas cilíndricas que lembram pilhas. Cada pilha
é um granum, que significa grào em latim. O
espaço interno do cloroplasto é preenchido por
um fluido viscoso chamado estroma, que corresponde à
matriz das mitocôndrias e contém DNA, enzimas
e ribossomos. Os cloroplastos são as centrais energéticas
da própria vida.
Funções
dos Plastos:
- Participação da Fotossíntese
(Cromoplastos);
- Armazenamento de Substâncias Nutritivas (Leucoplastos).
Nas células meristemáticas encontramos uma vesícula
primitiva denominada Proplasto, que na presença de
luz evolui para cromoplasto e na ausência de luz evolui
pra leucoplasto. Os proplastos são pequenas bolsas
esféricas, contendo em seu interior DNA, enzimas e
ribossomos, mas não há tilacóides e nem
clorofila. São capazes de se dividir e são herdados
de geração em geração celular.
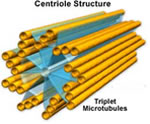
Centríolos
Os centríolos são estruturas citoplasmáticas
que estão presentes na maioria dos organismos eucariontes,
com exceção das plantas angiospermas (frutíferas).
O centríolo é um cilindro cuja parede é
constituída por nove conjuntos de três microtúbulos
e geralmente ocorrem aos pares nas células. Os centríolos
são desprovidos de membrana, são constituídos
por túbulos de natureza protéica (tubulina)
e recebem inúmeras denominações de acordo
com as funções que exercem como: diplossomos,
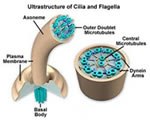
áster, cinetossomo, blefaroplastos, etc. Os centríolos
originam estruturas locomotoras denominadas cílios
e flagelos, que diferem entre si quanto ao comprimento e número
por célula e possuem um eixo de sustentação
chamado axonema (envolvido por uma membrana lipoprotéica).Os
flagelos são longos e pouco numerosos e executam ondulações
que se propagam da base em direção a extremidade
livre. Os cílios são curtos e muito numerosos
e executam um movimento semelhante ao de um chicote, com a
incrível freqüência de 10 a 40 batimentos
por segundo.
Funções
de Cílios e Flagelos:
- Locomoção da Célula;
- Movimentação de Líquido Extracelular;
- Limpeza das Vias Respiratórias.
Função dos Centríolos:
- Orientar a Divisão Celular,
pois originam uma estrutura denominada fuso mitótico,
onde se prendem os cromossomos;
- Originar Cílios e Flagelos.
Flagelos trabalham como chicotes que puxam
(como nas Chlamydomonas ou Halosphaera) ou empurrando (dinoflagellates,
um grupo de Protista unicelular) o organismo pela água.
Cílios trabalham como remos em um navio viking (o Paramecium
tem 17,000 cílios, cobrindo sua superfície exterior,
que remam dando-lhe movimento).
BIBLIOGRAFIA:
. BÚSSOLA ESCOLAR – www.bussolaescolar.com.br
|